|
奈良的成蛹阶段-Fbx15
千辛万苦脱了几层皮后,Shinya终于拥有了自己独立的实验室。第一次可以招帮手,好爽啊。但是问题又来了:研究生的生源是有限的,学生会倾向于选择资历更老条件更好的实验室,而不一定会选择刚起步的实验室;你想招但人家不来啊。为了吸引学生到他实验室,Shinya冥思苦想了好一阵,提出了一个雄心勃勃的计划,声称实验室的远景目标是研究怎么从终末分化的成体细胞变回多能的干细胞。
当时科学界的主流是研究怎么把胚胎多能干细胞分化成各种不同组织的细胞,以期用这些分化的功能细胞取代受损的或者有疾病的组织细胞。Shinya认为自己的实验室没有实力跟这些大牛竞争,那不如反其道而行之,研究怎么从分化的细胞逆转为多能干细胞。
当时科学界的主流观点认为,哺乳动物胚胎发育过程中的细胞分化是单向的,就像是时间不可逆转。这个观点也并非没有破绽,比如植物组织就具有多能性,一些植物的茎插入土壤会重新长出一棵植株,也即已经分化的茎细胞可以改变命运分化出新的根茎叶细胞。而早在1962年,也即Shinya出生的那一年,英国的John Gurdon爵士(与Shinya共享诺贝尔奖)报道了他的惊人发现:把蝌蚪的肠细胞核移植到去核的蛙卵中,新细胞可以发育成蝌蚪[7]。如果把杂合细胞发育到囊胚期,用囊胚期的细胞核再做一次核移植,那么就可以发育出可生育传代的成蛙[8]。进一步地,为了说服人们接受终末分化的细胞核也具有多能性,他把成蛙不同组织的细胞进行体外培养,发现核移植后来源不同的杂合细胞都可以发育到蝌蚪阶段[9]。1997年,Ian Wilmut和Keith Campbell基于同样的原理,把羊的乳腺细胞核移植到去核的羊卵中,成功地培育出了克隆羊多莉[10]。2001年,科学家发现,通过与 干细胞融合,胸腺细胞核获得了很大程度的重编程[11]。
Shinya计划的第一步是找到尽可能多的,类似于Nat1参与维持干细胞功能的因子(维持因子的意思是这些因子是胚胎干细胞在体外培养维持多能性所必需的)。他大胆推测,如果过表达这些维持因子也许可以让终末分化的细胞变回多能干细胞。一旦成功,诱导的多能干细胞会有着胚胎干细胞所不具备的优势:它不仅可以绕开胚胎干细胞引起的伦理问题,病人本身的诱导干细胞改造后重新植入病人时,由于是自身的细胞,将不会有免疫排斥的难题。
在这个远大前景的感召下,Shinya果然“忽悠”了三个学生加入他实验室。很快地,他们鉴定出一系列的在胚胎干细胞特异表达的基因。其中一个基因就是Fbx15。Shinya的学生Yoshimi Tokuzawa发现Fbx15除了特异表达于胚胎干细胞外,它还能被另外两个胚胎干细胞维持因子Oct3/4和Sox2直接调控。Shinya跟Yoshimi说:Fbx15应该参与维持干细胞多能性和胚胎的发育,我猜你没有办法得到Fbx15敲除的纯合鼠。Yoshimi构建质粒做了基因敲除小鼠,把染色体上的Fbx15基因通过同源重组替换成抗G418药物的基因neo。
复杂的生命又一次愚弄了Shinya:Fbx15敲除的纯合鼠活得很健康,没有显见的表型。Shinya又挑战他的学生说:好吧,Fbx15也许不是小鼠胚胎发育所必需的,但是它应该是维持体外胚胎干细胞所必需的,我打赌你没有办法在胚胎干细胞中彻底敲除这个基因。勤快的Yoshimi于是用较高浓度的G418从干细胞中筛到了纯合的敲除株,还是活得好好的,没有表型[12]。Shinya后来在回忆的时候打趣道:小鼠很happy,细胞也很happy,唯一不happy的就是可怜的学生Yoshimi了。
但是花这么多精力做的敲除小鼠不能就这么算了吧。Shinya又一次开动脑筋,想要废物利用。他发现由于Fbx15只在胚胎干细胞表达,Fbx15 promoter操控的抗药基因neo在成体的成纤维细胞里不表达,所以细胞对药物 G418敏感;而敲除鼠里得到的胚胎干细胞却可以在很高浓度的 G418中生长。如果终末分化的成纤维细胞能诱导成胚胎干细胞,那么它就会产生对G418的 抗药性。即便成纤维细胞只是获得了部分胚胎干细胞的特性,那么它也应该能抗低浓度的 G418 (图二)。Fbx15敲除鼠实际上提供了很好的筛选诱导干细胞的系统!
京都大学的化蛹成蝶阶段-iPS
凭借他鉴定胚胎干细胞维持因子的出色工作,2004年Shinya在名气更大的京都大学找到新的职位。除了Fbx15敲除鼠的筛选系统,Shinya还积累了他鉴定的加上文献报道的24个维持因子。Shinya跃跃欲试,他准备破壳而出,拍翅成蝶了!
Shinya的另一位学生Kazutoshi Takahashi此前已经发表了一篇关于干细胞致癌性的Nature文章。Shinya决意让他来承担最大胆的课题-逆分化成体细胞,因为他知道,有一篇Nature文章保底,即便接下来的几年一无所获,他的学生也能承受得了。
http://image.sciencenet.cn/album/201210/14/141853dewtw5036bwdbwm5.jpg
图二:筛选iPS的系统。在Fbx15敲除鼠基因组,Fbx15 基因被bgeo基因(β-galactosidase 和neo的融合基因)取代。成鼠角形的成纤维细胞中,内源的Fbx15 promoter关闭,bgeo不能表达,细胞在G418药物处理下会死亡;在圆形的多能干细胞中,Fbx15 promoter会启动bgeo,细胞能在G418中生长。如果成纤维细胞感染携带干细胞维持因子的逆转录病毒,并能够被逆分化成干细胞,那么它就能逃过G418的选择压力,增殖形成细胞克隆[13]。
即便有很好的筛选系统,这个课题在当初看来也是非常冒险甚至是不可行的。当时的人们普遍认为成体细胞失去了多能性,也许成体细胞本身就是不可逆转的,你做什么也没有用。即便通过转核技术实现了成体细胞核命运的逆转,那也只是细胞核,不是整个细胞。胚胎细胞和成体细胞的染色体是一样的,细胞核具有全能性,尚可理解。而且要实现细胞核的逆转还需要转到卵细胞,让卵细胞质帮助它重编程,而卵细胞质中的蛋白不计其数。如果要实现整个细胞命运的逆转需要让细胞质中所有的蛋白重新洗牌。即便细胞可以重新编程,那也应该是很多蛋白共同参与的。Shinya当年在手上的仅仅是24个因子。也许有另外几百几千种因子被遗漏,缺少其中一种都无法实现重编程。用这24个因子异想天开要实现细胞重编程,根据已有的知识从逻辑上讲可能性几乎为零。
Kazutoshi这个愣头青不管这些,他给成纤维细胞一一感染过表达这些因子的病毒,结果当然没有筛选到任何抗 G418的细胞。Shinya知道如何保持学生的斗志,他故作镇定地说:你看,这说明我们的筛选系统很好啊,没有出现任何假阳性。
在试了一遍无果后,Kazutoshi大胆提出想把24个病毒混合起来同时感染细胞。Shinya觉得这是很愚蠢的想法:没人这么干过啊同学,不过死马当作活马医,你不嫌累的话就去试吧。
等了几天,奇迹竟然发生了。培养板上稀稀疏疏地竟然出现了十几个抗 G418的细胞克隆!一个划时代的发现诞生了。
关键实验取得突破以后,其后的事情就按部就班了。Kazutoshi每次去掉一个病毒,把剩下的23个病毒混合感染成体细胞,看能长多少克隆,以此来鉴别出哪一些因子是诱导干细胞所必需的。最后他鉴定出了四个明星因子:Oct3/4, Sox2, c-Myc,和 Klf4。这四个因子在成纤维细胞中过表达,就足以把它逆转为多能干细胞!
那抗 G418的细胞克隆就一定是多能干细胞吗?他们通过一系列的指标,比如基因表达谱,分化潜能等,发现这些细胞在相当大的程度上与胚胎干细胞相似。
2006年Shinya报道了小鼠诱导干细胞,引起科学界轰动[13];2007年,他在人的细胞中同样实现了细胞命运的逆转,科学界沸腾了[14]。
展望
回过头来,种种不可能,Shinya怎么就幸运地成功了呢?现在通过更多的研究,我们知道,干细胞特性的维持是由一个基因网络来共同作用的,通过上调某些关键基因就可以重建这个网络,逆转细胞的命运;山中伸弥最后鉴定的四个因子也不是必须的,用24个因子以外的其它因子进行组合可以达到同样的目的。这好比是一张大网,你只要能撑起其中的几个支点,就可以把整张网撑起来。
iPS的发现有着不同寻常的意义。首先,它更新了人们的观念,从此之后人们不再认为细胞的命运不可逆转,不单可以逆转,细胞其实还可以实现不同组织间的转分化(Transdifferentiation)
。其次,iPS细胞绕过了胚胎干细胞的伦理困境,很多实验室都可以重复这个简单的实验得到iPS,开展多能干细胞的研究。其三,iPS细胞具有很多胚胎干细胞所没有的优势:来自于病人自身的iPS细胞体外操作后重新植入病人体内,免疫反应将大大减少;如果将病人的体细胞逆转为ips细胞,在体外分化观察在这个过程中出现的问题,就可以实现在培养皿里某种程度上模拟疾病的发生;疾病特异的iPS在体外扩增和分化以后,还可以用于筛选治疗该疾病的药物,或者对药物的毒性进行检测(图三)。
http://image.sciencenet.cn/album/201210/14/141853i545hi4mk5dshmiz.jpg
图三:iPS细胞的潜在用途。采自病人的少量成体细胞被逆分化成iPS细胞后,能够在体外增殖,改造,分化成组织特异性的功能细胞。这些功能细胞重新植入人体可以帮助/取代受损的或者得病的器官/组织。iPS或者这些功能细胞也可以作为疾病模型用于一些药物的筛选和毒性测试(图片来自诺贝尔奖网站)。
但是这仅仅是新的开始,生命科学如此复杂和不可预测,要把这些愿景变成现实,让iPS真正造福人类,这其中还有重重的困难。Shinya Yamanka,这位科学的宠儿,怀着最初帮助更多病人的理想,无畏地踏上了新的征程。 | 



 狗仔卡
狗仔卡
 发表于 2015-1-21 16:38:06
发表于 2015-1-21 16:38:06
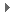
 收藏
收藏 顶
顶 踩
踩 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 显身卡
显身卡 楼主
楼主